
8
Оказалось, что почти все об этом генераторе выяснил президент Королевского общества Англии Алан Ходжкин. Его работы были настоящим успехом физики в биологии (Hodgkin, 1937), за что он и был удостоен Нобелевской премии.
В советской физиологии в то время господствовала фазовая теория. Было ясно, что изменение знака потенциала при генерации нервного импульса фазовыми свойствами протоплазмы не объяснишь. И я придумал не противоречащую уже поставленным экспериментам фазово-мембранную гипотезу (Либерман Е.А., 1961). Сторонник фазовой теории А.С. Трошин включил изложение этой гипотезы в свой доклад в Англии и в свою книжку. Он говорил, что именно этот доклад вызвал эксперименты Алена Ходжкина с перфузией гигантского аксона кальмара, которые убили фазово-мембранную гипотезу наповал: обычные нервные импульсы генерировала мембрана, лишенная протоплазмы. К счастью, у меня появились замечательные сотрудники Алексей Владимирович Бабаков и Лев Николаевич Ермишкин, и мы начали моделировать нервный импульс на искусственных фосфолипидных мембранах (Babakov et al., 1966).
Нервно-мышечный синапс
Английские ученые Поль Фэтт и Бернард Катц обнаружили, что медиатор выделяется из нервных окончаний порциями (Fatt, Katz, 1952). В покое каждая порция вызывает на мембране мышечного волокна маленькое изменение разности потенциалов, которое так и назвали «миниатюрным потенциалом». Порции ацетилхолина спонтанно выделяются в покое. Нервный импульс в тысячи раз увеличивает среднюю частоту их выделения. Множество одновременно возникающих миниатюрных потенциалов сливаются и образуют синаптический потенциал. Когда мы с помощью все той же установки для счета альфа-частиц научились отводить миниатюрные потенциалы от нервно-мышечного соединения лягушки, оставалось решить вопрос о том, почему приходящий нервный импульс учащает слипание синаптических пузырьков с наружной пресинаптической мембраной нервного окончания.
Сначала казалось, что причиной является уменьшение разности потенциалов на мембране, вызванное приходом нервного импульса.
Действительно, в нормальном солевом растворе всевозможные воздействия, уменьшающие эту разность потенциалов, увеличивают частоту миниатюрных потенциалов. Но если из наружного раствора удалить ионы кальция, то увеличения частоты не происходит, несмотря на приход нервного импульса.
Но самым непонятным было резкое увеличение частоты выделения квантов при повышении концентрации любых непроникающих ионов или нейтральных молекул, например, сахарозы в наружном солевом растворе. Жанне Львовне Блиох, Ирине Михайловне Глаголевой и мне (Blioch et al., 1968) удалось разобраться в этой запутанной ситуации. У нас был удобный для изучения миниатюрных потенциалов нервно-мышечный препарат грудной мышцы лягушки, управляющей движением кожи. Этот препарат состоял, в основном, из одного слоя мышечных волокон, к которым нерв подходил перпендикулярно и разветвлялся в виде причудливого дерева, посылая нервные окончания к одиночным мышечным волокнам. На простой парафиновой пластинке, на которой обычно прикрепляется нервно-мышечный препарат лягушки, прозрачных волокон этой грудной мышцы почти не было видно. Но по совету Лилии Мироновны Цофиной, которая нашла для нас этот препарат, добавка к парафину растертого активированного угля позволила отлично видеть мышечные волокна и нервные окончания. На этом препарате удалось получать миниатюрные потенциалы, качество которых оценил при посещении нашей лаборатории сам великий Бернард Катц.
Мы исходили из простой физической идеи: для слипания пузырьков с наружной мембраной пресинаптического волокна надо уменьшить ее поверхностный электрический заряд.
Это могут делать ионы кальция. Опыты по слипанию искусственных фосфолипидных мембран ставил Валерий Аркадиевич Ненашев. В растворах с кальцием слипание таких мембран происходит гораздо быстрей.
9
Каждый нервный импульс вызывает не только деполяризацию пресинаптического окончания, но и вход внутрь окончания нервного волокна ионов кальция, рассуждали мы. Поэтому в растворе без кальция нет синаптической передачи. Одновременно эта гипотеза объясняла зависимость частоты миниатюрных потенциалов от осмотического давления. Этот эффект долго не находил объяснения, поскольку осмотическое давление никак не влияет на распространение нервного импульса. Идея о снижении поверхностного заряда сразу объяснила влияние осмотического давления: повышение концентрации ионов или нейтральных молекул в наружном растворе приводит к тому, что из клетки выходит вода и, следовательно, увеличивается внутриклеточная концентрация катионов, в том числе и кальция.
Мы послали статью Бернарду Катцу, который до того посетил нас и любовался своими миниатюрными потенциалами. Статью задержали надолго. Поль Фэтт перевел ее на настоящий английский. Пока наша статья была у них, Бернард Катц сообщил, что удалось поставить прямые опыты с введением ионов Са++ в гигантские синапсы кальмаров. У этих синапсов толщина пресинаптического волокна достигает миллиметра.
Он попросил нас сослаться на эти опыты, которые, по моему мнению, нельзя воспроизвести, поскольку кальций на пути к наружной мембране поглотят митохондрии. Добавляя разобщители окислительного фосфорилирования, Ирина Михайловна Глаголева и Заур Хаджи-Мурадович Хашаев сумели освобождать кальций внутри нервного окончания из митохондрий.
Оказалось, что все вещества, снимающие электрическое поле на мембране митохондрий, резко повышают частоту выделения квантов ацетилхолина (Глаголева И.М. и др., 1970).
Электрическая энергетика живой клетки
На семинаре И.М. Гельфанда Владимир Скулачев рассказал, что в биоэнергетике появился англичанин Питер Митчелл, которого не понимает никто, и дал ссылку на его статью (Mitchell, 1961). Мне повезло, что я плохо знаю английский, иначе меня всегда подавлял бы поток научной литературы. Статью переводила Лилия Мироновна Цофина, с которой я работаю около пятидесяти лет. Цофина окончила кафедру биохимии животных биофака МГУ, и с ней мы разбирались в малопонятных тогда новых идеях Митчелла о важных свойствах митохондриальной мембраны. После первых фраз статьи я подумал, что дальше пойдет речь о разности потенциалов на мембране митохондрий. Однако, автор думал, что возникнет лишь градиент pH. Он не знал тогда работ Алена Ходжкина. Идею Питера Митчелла о том, что разобщители окислительного фосфорилирования являются переносчиками протонов, мы начали проверять вместе со Львом Исаевичем Кришталиком, Вячеславом Семеновичем Маркиным и Василием Павловичем Топалы (Liberman and Topaly, 1968). Одновременно вместе с Лилией Мироновной Цофиной мы измеряли мембранный потенциал митохондрий (Рис.2)(Либерман Е.А., Цофина Л.М., 1969). Лилия Цофина, Пулат Исаев и я измерили также мембранный потенциал фоточастиц бактерий (Рис.3) (Isaev et al., 1970).
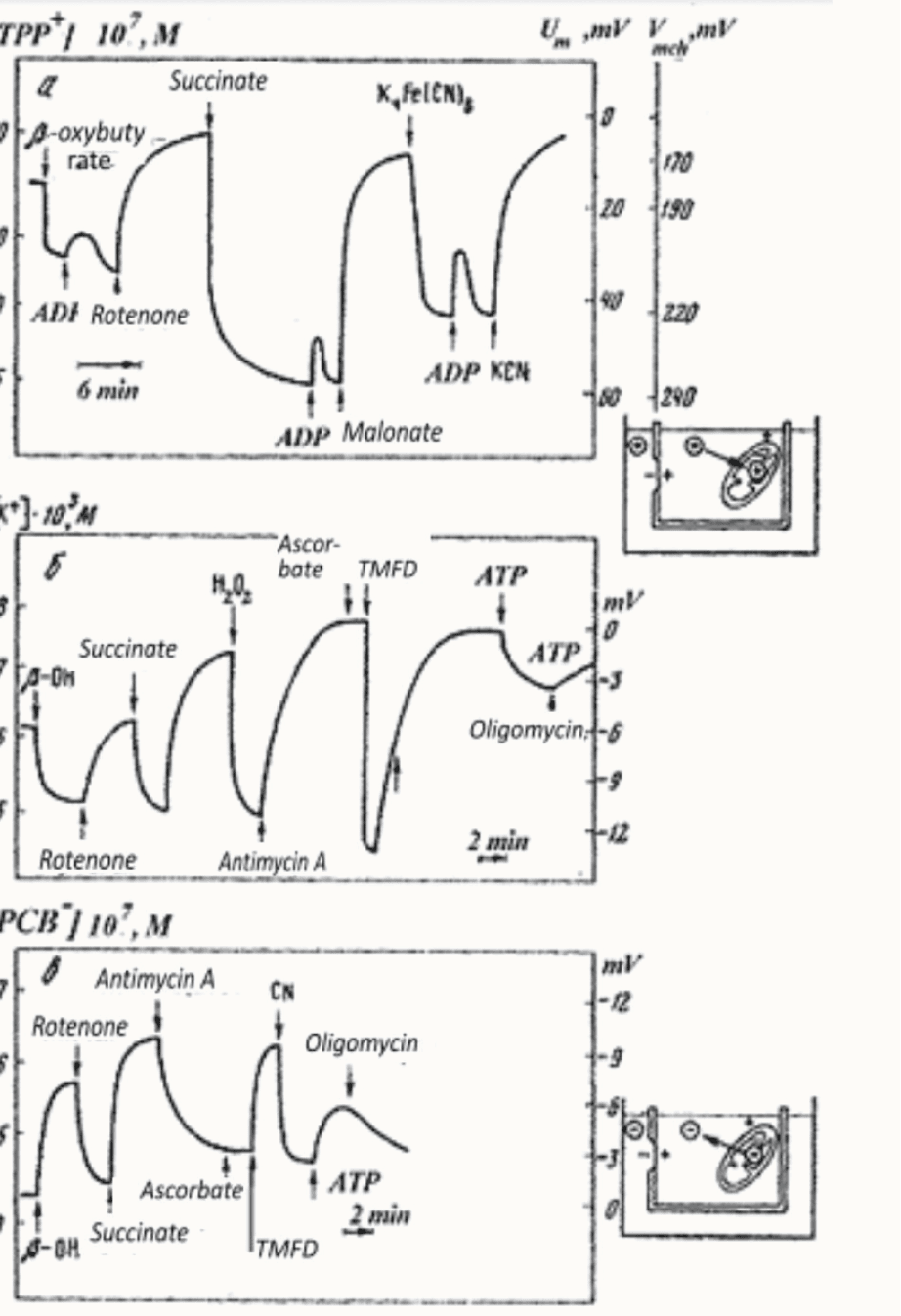
Рис. 2. Изучение мембранного потенциала митохондрий.
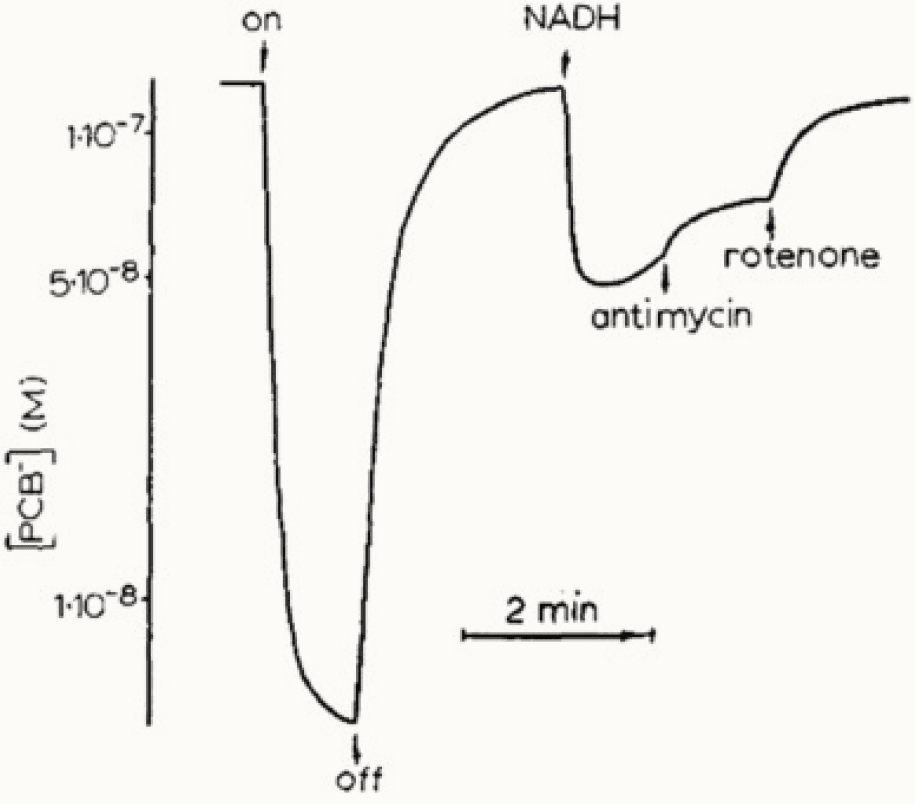
Рис. 3. Транспорт ФКБ в хроматофорах фотосинтезирующих бактерий. На оси ординат — концентрация проникающих ионов.
Все эти измерения велись методом проникающих ионов, разработанным В.П. Топалы и мной (Либерман Е.А., Топалы В.П., 1969).
Мы работали на митохондриях, которые специально для нас выделяли Елена Александровна Пинус и Инна Алексеевна Горская. Субмитохондриальные частицы делал Валдур Сакс, а фоточастицы — Виктор Дмитриевич Самуилов. Рабочая атмосфера в молекулярном корпусе биофака МГУ, где также выполнялись эти работы, создавалась талантливым организатором Владимиром Петровичем Скулачевым, который активно участвовал в исследованиях, сомневался и держал нас в курсе достижений мировой науки.
Как в мембране митохондрий работает протонный канал, выяснила Марина Алексеевна Владимирова. Она сама выделяла субмитохондриальные частицы, сама сдирала с них АТФазу и сама измеряла, как меняется мембранный потенциал после добавления олигомицина. Эксперимент доказал, что этот ингибитор действительно закрывает протонный канал АТФазы (Grinius et al., 1970).
За серию работ по экспериментальному доказательству новой роли белков мембраны митохондрий в переносе зарядов в процессе синтеза АТФ в 1975 году мне, В.П. Скулачеву, Л.М. Цофиной и А. Ясайтису была присуждена Госпремия СССР.
Однако вскоре моя роль в биоэнергетике сошла на нет благодаря появлению среди нас настоящего экспериментатора Леля Александровича Драчева (Drachev et al., 1974). Пришлось продолжить путь к Хаиматике. На этом пути меня все время поддерживал и служил “рабочей лошадкой” Анатолий Яковлевич Зильберштейн.


Ефим Либерман (снизу слева), Светлана Минина (снизу справа), Василий Топалы (сверху слева), Анатолий Зильберштейн (сверху справа), 1970-е годы.
10
хаиматика
хаиматика
Итогом жизни в науке стало установление связей между биологией, физикой, математикой и новая область исследования, посвященная вычислениям в живых системах. Ефим Либерман дал имя новой науке: «Хаиматика»
I
ДНК – это текст программы для молекулярных компьютеров клеток. «Текст» по определению не случайная последовательность знаков и может существовать только внутри языковой системы. В данном случае это генетический язык, изоморфный естественному языку
II
Вычисление в живой клетке является реальным физическим действием и требует затрат свободной энергии и времени. Поскольку все живые организмы состоят из клеток, это относится ко всему управлению, которое осуществляется в биосфере
III
Молекулярные вычисления ограничены микроскопическим объемом клетки и принципиальной возможностью влияния вычисления на условия решаемой задачи: квантовая механика возникла из осознания реальности измерения, Хаиматика - из реальности вычисления
IV
Для решения сложных задач клетка создает устройство квантового вычисления, использующего кванты гиперзвука и клеточный цитоскелет, как вычисляющую среду. Цена вычисления в таком компьютере стремится к физическому пределу – постоянной Планка
Утверждения Хаиматики просты, но они требуют изменения традиционных представлений, принятых в научной практике
Читать книгу
Глава I
Как все начиналось
хаиматика